Early human migrations
Definition
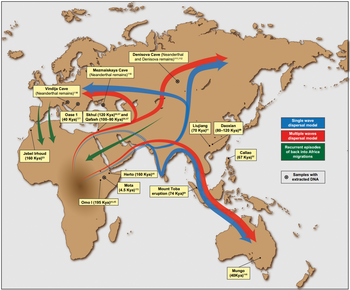
Early human migrations are the earliest migrations and expansions of archaic and modern humans across continents beginning 2 million years ago with the out of Africa migration of Homo erectus. This initial migration was followed by other archaic humans including H. heidelbergensis, which lived around 500,000 years ago and was the likely ancestor of both Denisovans and Neanderthals.
Within Africa, Homo sapiens dispersed around the time of its speciation, roughly 300,000 years ago. The "recent African origin" paradigm suggests that the anatomically modern humans outside of Africa descend from a population of Homo sapiens migrating from East Africa roughly 70,000 years ago and spreading along the southern coast of Asia and to Oceania before 50,000 years ago. Modern humans spread across Europe about 40,000 years ago.
The migrating modern human populations are known to have interbred with local varieties of archaic humans, so that contemporary human populations are descended in small part (below 10% contribution) from regional varieties of archaic humans.
After the Last Glacial Maximum, North Eurasian populations migrated to the Americas about 20,000 years ago. Northern Eurasia was peopled after 12,000 years ago, in the beginning Holocene. Arctic Canada and Greenland were reached by the Paleo-Eskimo expansion around 4,000 years ago. Finally, Polynesia was peopled after 2,000 years ago, by the Austronesian expansion.
Early humans (before Homo sapiens)

The earliest humans developed out of australopithecine ancestors after about 3 million years ago, most likely in Eastern Africa, most likely in the area of the Kenyan Rift Valley, where the oldest known stone tools were found. Stone tools recently discovered at the Shangchen site in China and dated to 2.12 million years ago are claimed to be the earliest known evidence of hominins outside Africa, surpassing Dmanisi in Georgia by 300,000 years.
Homo erectus
Between 3 and 2 million years ago, Homo erectus spread throughout East Africa and to Southern Africa (Telanthropus capensis), but not yet to West Africa. Around 1.9 million years ago, Homo erectus migrated out of Africa via the Levantine corridor and Horn of Africa to Eurasia. This migration has been proposed as being related to the operation of the Saharan pump, around 1.9 million years ago. Homo erectus dispersed throughout most of the Old World, reaching as far as Southeast Asia. Its distribution is traced by the Oldowan lithic industry, by 1.3 million years ago extending as far north as the 40th parallel (Xiaochangliang), and its late phase (after 0.5 million years ago) as far as the 47th parallel (Vértesszőlős) in Europe.
Key sites for this early migration out of Africa are Riwat in Pakistan (~2 Ma?), Ubeidiya in the Levant (1.5 Ma) and Dmanisi in the Caucasus (1.81 ± 0.03 Ma, p = 0.05).
China was populated as early as 1.66 Mya based on stone artifacts found in the Nihewan Basin. The archaeological site of Xihoudu (西侯渡) in Shanxi Province is the earliest recorded use of fire by Homo erectus, which is dated 1.27 million years ago.
Southeast Asia (Java) was reached about 1.7 million years ago (Meganthropus). Western Europe was first populated around 1.2 million years ago (Atapuerca).
Robert G. Bednarik has suggested that Homo erectus may have built rafts and sailed oceans, a theory that has raised some controversy.
After H. erectus

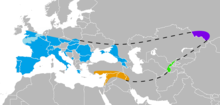
One million years after its dispersal, H. erectus was diverging into new species. H. erectus is a chronospecies and was never extinct, so that its "late survival" is a matter of taxonomic convention. Late forms of H. erectus are thought to have survived until after about 0.5 million ago, with derived forms classified as H. antecessor in Europe around 800,000 years ago and H. heidelbergensis in Africa around 600,000 years ago. H. heidelbergensis in its turn spreads across East Africa (H. rhodesiensis) and to Eurasia, where it gives rise to Neanderthals and Denisovans.
H. heidelbergensis, Neanderthals and Denisovans expanded north beyond the 50th parallel (Eartham Pit, Boxgrove 500kya, Swanscombe Heritage Park 400kya, Denisova Cave 50 kya). It has been suggested that late Neanderthals may even have reached the boundary of the Arctic, by c. 32,000 years ago, when they were being displaced from their earlier habitats by H. sapiens, based on 2011 excavations at the site of Byzovaya in the Urals (Komi Republic,
65.02°N 57.42°E).
65.02°N 57.42°E).
Other archaic human species are assumed to have spread throughout Africa by this time, although the fossil record is sparse. Their presence is assumed based on traces of admixture with modern humans found in the genome of populations indigenous to Southern and West Africa. Homo naledi, discovered in South Africa in 2013 and tentatively dated to about 300,000 years ago, may represent fossil evidence of such an archaic human species.
Neanderthals spread across the Near East and Europe, while Denisovans appear to have spread across Central and East Asia and to Southeast Asia and Oceania. There is evidence that Denisovans interbred with Neanderthals in Central Asia where their habitats overlapped.
It is most likely from an African variety of H. antecessor that H. sapiens develops around 300,000 years ago.
Homo sapiens

Dispersal throughout Africa
Homo sapiens (anatomically modern humans) are assumed to have emerged about 300,000 years ago based on thermoluminescence dating of artefacts from Jebel Irhoud, Morocco, published in 2017. Previously, the Omo remains, excavated between 1967 and 1974 in Omo National Park, Ethiopia, and dated to 200,000 years ago, were long held to be the oldest known fossils of anatomically modern humans.
Early modern humans expanded to Western Eurasia, Central, Western and Southern Africa from the time of their emergence. While early expansions to Eurasia appear not to have persisted, expansions to Southern and Central Africa resulted in the deepest temporal divergence in living human populations. Early modern human expansion in sub-Saharan Africa appears to contribute to the end of late Acheulean (Fauresmith) industries at about 130,000 years ago, although very late coexistence of archaic and early modern humans, until as late as 12,000 years ago, has been argued for West Africa in particular.
The ancestors of the modern Khoi-San expanded to Southern Africa before 150,000 years ago, possibly as early as before 260,000 years ago, so that by the beginning of the MIS 5 "megadrought", 130,000 years ago, there were two ancestral population clusters in Africa, bearers of mt-DNA haplogroup L0in southern Africa, ancestral to the Khoi-San, and bearers of haplogroup L1-6 in central/eastern Africa, ancestral to everyone else. There was a significant back-migration of bearers of L0 towards eastern Africa between 120 and 75 kya.
Expansion to Central Africa, by the ancestors of the Central African forager populations (African Pygmies) most likely took place before 130,000 years ago, and certainly before 60,000 years ago.
The situation in West Africa is difficult to interpret due to a sparsity of fossil evidence. Homo sapiens seems to have reached the western Sahelian zone by 130 kya, while tropical West African sites associated with H. sapiens are known only from after 130 kya. Unlike elsewhere in Africa, archaic MSA sites appear to persist until very late, down to the Holocene boundary (12 kya), pointing to the possibility of late survival of archaic humans, and late hybridization with H. sapiens in West Africa.
Early northern Africa dispersal
Populations of H. sapiens migrated to the Levant and to Europe between 130,000 and 115,000 years ago, and possibly in earlier waves as early as 185,000 years ago. These early migrations do not appear to have led to lasting colonisation and receded by about 80,000 years ago. There is a possibility that this first wave of expansion may have reached China (or even to North America) as early as 125,000 years ago, but would have died out without leaving a trace in the genome of contemporary humans.
There is some evidence for the argument that modern humans left Africa at least 125,000 years ago using two different routes: through the Nile Valley heading to the Middle East, at least into modern Israel (Qafzeh: 120,000–100,000 years ago); and a second one through the present-day Bab-el-Mandeb Strait on the Red Sea (at that time, with a much lower sea level and narrower extension), crossing it into the Arabian Peninsula, settling in places like the present-day United Arab Emirates (125,000 years ago) and Oman (106,000 years ago) and then possibly going into the Indian Subcontinent (Jwalapuram: 75,000 years ago). Despite the fact that no human remains have yet been found in these three places, the apparent similarities between the stone tools found at Jebel Faya, the ones from Jwalapuram and some African ones suggest that their creators were all modern humans. These findings might give some support to the claim that modern humans from Africa arrived at southern China about 100,000 years ago (Zhiren Cave, Zhirendong, Chongzuo City: 100,000 years ago; and the Liujiang hominid (Liujiang County): controversially dated at 139,000–111,000 years ago ). Dating results of the Lunadong (Bubing Basin, Guangxi, southern China) teeth, which include a right upper second molar and a left lower second molar, indicate that the molars may be as old as 126,000 years.
Since these previous exits from Africa did not leave traces in the results of genetic analyses based on the Y chromosome and on MtDNA (which represent only a small part of the human genetic material), it seems that those modern humans did not survive or survived in small numbers and were assimilated by our major antecessors. An explanation for their extinction (or small genetic imprint) may be the Toba catastrophe theory (74,000 years ago). However, some argue that its impact on the human population was not dramatic.
An Asia center of origin and dispersal for the mtDNA haplogroup L3 has also been hypothesized based on the fossil record, the similar coalescence dates of L3 and its Eurasian-distributed M and N derivative clades (~71 kya), the distant location in Southeast Asia of the oldest subclades of M and N, and the comparable age of the paternal haplogroup DE. After an initial Out-of-Africa migration of early anatomically modern humans around 125 kya, fully modern human L3-carrying females are thus proposed to have back-migrated from the maternal haplogroup's place of origin in Eurasia around 70 kya along with males bearing the paternal haplogroup E, which is also thought to have originated in Eurasia. These new Eurasian lineages are then suggested to have largely replaced the old autochthonous male and female African lineages.
Coastal migration
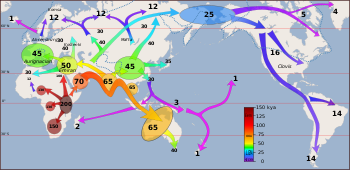
The so-called "recent dispersal" of modern humans has taken place after beginning about 70,000 years ago. It is this migration wave that led to the lasting spread of modern humans throughout the world.
A small group of members of a population inhabiting East Africa, who were bearers of the mitochondrial haplogroup L3 and numbered possibly fewer than 1,000 individuals, crossed the Red Sea strait at Bab el Mandib, to what is now Yemen, after around 75,000 years ago. A recent review has shown support for both the Northern Route through Sinai/Israel/Syria (Levant), and, that both routes may have been used. Their descendants spread along the coastal route around Arabia and Persia to the Indian subcontinent before 55,000 years ago. The coastal migration scenario between roughly 70,000 and 50,000 years ago is associated with mitochondrial haplogroups M and N, both derivative of L3.
A fragment of a jawbone with eight teeth found at Misliya Cave, Israel, have been dated to around 185,000 years ago. Layers dating from between 250,000 and 140,000 years ago in the same cave contained tools of the Levallois type which could put the date of the first migration even earlier if the tools can be associated with the modern human jawbone finds.
Along the way H. sapiens interbred with Neanderthals and Denisovans, with Denisovan DNA making 0.2% of mainland Asian and Native American DNA.
Oceania
Migrations continued along the Asian coast to Southeast Asia and Oceania, colonising Australia before 50,000 years ago. By reaching Australia, H. sapiens for the first time expanded its habitat beyond that of H. erectus.
Denisovan ancestry is shared by Melanesians, Australian Aborigines, and smaller scattered groups of people in Southeast Asia, such as the Mamanwa, a Negrito people in the Philippines suggesting the interbreeding took place in Eastern Asia where the Denisovans lived. Denisovans may have crossed the Wallace Line, with Wallacea serving as their last refugium. Homo erectus crossed the Lombok gap reaching as far as Flores, but never made it to Australia.
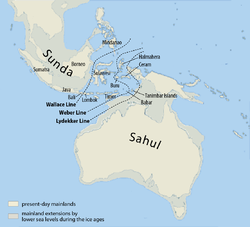
During this time sea level was much lower and most of Maritime Southeast Asia formed one land mass known as Sunda. Migration continued Southeast on the coastal route to the straits between Sunda and Sahul, the continental land mass of present-day Australia and New Guinea. The gaps on the Weber Line are up to 90 km wide, so the migration to Australia and New Guinea would have required seafaring skills. Migration also continued along the coast eventually turning northeast to China and finally reaching Japan before turning inland. This is evidenced by the pattern of mitochondrial haplogroups descended from haplogroup M, and in Y-chromosome haplogroup C.
Sequencing of one Aboriginal genome from an old hair sample in Western Australia, revealed that the individual was descended from people who migrated into East Asia between 62,000 and 75,000 years ago. This supports the theory of a single migration into Australia and New Guinea before the arrival of Modern Asians (between 25,000 and 38,000 years ago) and their later migration into North America. This migration is believed to have happened around 50,000 years ago, before Australia and New Guinea were separated by rising sea levels approximately 8,000 years ago. This is supported by a date of 50,000 - 60,000 years ago for the oldest evidence of settlement in Australia, around 40,000 years ago for the oldest human remains, the earliest humans artifacts which are at least 65,000 years old and the extinction of the Australian megafauna by humans between 46,000 and 15,000 years ago argued by Tim Flannery, which is similar to what happened in the Americas. The continued use of stone age tools in Australia has been much debated.
Dispersal throughout Eurasia
The population brought to South Asia by coastal migration appears to have remained there for some time, during roughly 60,000 to 50,000 years ago, before spreading further throughout Eurasia. This dispersal of early humans, at the beginning of the Upper Paleolithic, gave rise to the major population groups of the Old World and the Americas.
Towards the West, Upper Paleolithic populations associated with mitochondrial haplogroup R and its derivatives, spread throughout Asia and Europe, with a back-migration of M1 to North Africa and the Horn of Africa several millennia ago.
Presence in Europe is certain after 40,000 years ago, possibly as early as 43,000 years ago, rapidly replacing the Neanderthal population. Contemporary Europeans have Neanderthal ancestry, but it seems likely that substantial interbreeding with Neanderthals ceased before 47,000 years ago, i.e. took place before modern humans entered Europe.
There is evidence from mitochondrial DNA that modern humans have passed through at least one genetic bottleneck, in which genome diversity was drastically reduced. Henry Harpending has proposed that humans spread from a geographically restricted area about 100,000 years ago, the passage through the geographic bottleneck and then with a dramatic growth amongst geographically dispersed populations about 50,000 years ago, beginning first in Africa and thence spreading elsewhere. Climatological and geological evidence suggests evidence for the bottleneck. The explosion of Lake Toba created a 1,000 year cold period, as a result of the largest volcanic eruption of the Quaternary, potentially reducing human populations to a few tropical refugia. It has been estimated that as few as 15,000 humans survived. In such circumstances genetic drift and founder effects may have been maximised. The greater diversity amongst African genomes may be in part due to the greater prevalence of African refugia during the Toba incident. However, a recent review highlights that the single-source hypothesis of non-African populations is less supported by ancient DNA analysis than multiple sources plus genetic mixing across Eurasia.
Europe
The recent expansion of anatomically modern humans reached Europe around 40,000 years ago, from Central Asia and the Middle East, as a result of cultural adaption to big game hunting of sub-glacial steppe fauna. Neanderthals were present both in the Middle East and in Europe, and the arriving populations of anatomically modern humans (also known as "Cro-Magnon" or European early modern humans) have interbred with Neanderthal populations to a limited degree. Populations of modern humans and Neanderthal overlapped in various regions such as in Iberian peninsula and in the Middle East. Interbreeding may have contributed Neanderthal genes to palaeolithic and ultimately modern Eurasians and Oceanians.
An important difference between Europe and other parts of the inhabited world was the northern latitude. Archaeological evidence suggests humans, whether Neanderthal or Cro-Magnon, reached sites in Arctic Russia by 40,000 years ago.
Cro-Magnon are considered the first anatomically modern humans in Europe. They entered Eurasia by the Zagros Mountains (near present-day Iran and eastern Turkey) around 50,000 years ago, with one group rapidly settling coastal areas around the Indian Ocean and one group migrating north to steppes of Central Asia. Modern human remains dating to 43-45,000 years ago have been discovered in Italy and Britain, with the remains found of those that reached the European Russian Arctic 40,000 years ago.
Humans colonised the environment west of the Urals, hunting reindeer especially, but were faced with adaptive challenges; winter temperatures averaged from −20 to −30 °C (−4 to −22 °F) while fuel and shelter were scarce. They travelled on foot and relied on hunting highly mobile herds for food. These challenges were overcome through technological innovations: production of tailored clothing from the pelts of fur-bearing animals; construction of shelters with hearths using bones as fuel; and digging of “ice cellars” into the permafrost for storing meat and bones.
A mitochondrial DNA sequence of two Cro-Magnons from the Paglicci Cave in Italy, dated to 23,000 and 24,000 years old (Paglicci 52 and 12), identified the mtDNA as Haplogroup N, typical of the latter group.



The expansion of modern human population is thought to have begun 45,000 years ago, and may have taken 15,000-20,000 years for Europe to be colonized.
During this time the Neanderthals were slowly being displaced. Because it took so long for Europe to be occupied, it appears that humans and Neanderthals may have been constantly competing for territory. The Neanderthals had larger brains, and were larger overall, with a more robust or heavily built frame, which suggests that they were physically stronger than modern Homo sapiens. Having lived in Europe for 200,000 years, they would have been better adapted to the cold weather. The anatomically modern humans known as the Cro-Magnons, with widespread trade networks, superior technology and bodies likely better suited to running, would eventually completely displace the Neanderthals, whose last refuge was in the Iberian peninsula. After about 25,000 years ago the fossil record of the Neanderthals ends, indicating that they had become extinct. The last known population lived around a cave system on the remote south-facing coast of Gibraltar from 30,000 to 24,000 years ago.
From the extent of linkage disequilibrium, it was estimated that the last Neanderthal gene flow into early ancestors of Europeans occurred 47,000–65,000 years BP. In conjunction with archaeological and fossil evidence, the gene flow is thought likely to have occurred somewhere in Western Eurasia, possibly the Middle East. Studies show a higher Neanderthal admixture in East Asians than in Europeans.North African groups share a similar excess of derived alleles with Neanderthals as do non-African populations, whereas Sub-Saharan African groups are the only modern human populations that generally did not experience Neanderthal admixture. The Neanderthal-linked haplotype B006 of the dystrophin gene has also been found among nomad pastoralist groups in the Sahel and Horn of Africa, who are associated with northern populations. Consequently, the presence of this B006 haplotype on the northern and northeastern perimeter of Sub-Saharan Africa is attributed to gene flow from a non-African point of origin.
East and North Asia
"Tianyuan Man", an individual who lived in China c. 40,000 years ago, showed substantial Neanderthal admixture. A 2017 study of the ancient DNA of Tianyuan Man found that the individual is closely related to modern East Asian populations, but not a direct ancestor. A 2013 study found Neanderthal introgression of 18 genes within the chromosome 3p21.31 region (HYAL region) of East Asians. The introgressive haplotypes were positively selected in only East Asian populations, rising steadily from 45,000 years ago until a sudden increase of growth rate around 5,000 to 3,500 years ago. They occur at very high frequencies among East Asian populations in contrast to other Eurasian populations (e.g. European and South Asian populations). The findings also suggests that this Neanderthal introgression occurred within the ancestral population shared by East Asians and Native Americans.
A 2016 study presented an analysis of the population genetics of the Ainu people of northern Japan as key to the reconstruction of the early peopling of East Asia. The Ainu were found to represent a more basal branch than the modern farming populations of East Asia, suggesting an ancient (pre-Neolithic) connection with northeast Siberians. A 2013 study associated several phenotypical traits associated with Mongoloids with a single mutation of the EDAR gene, dated to c. 35,000 years ago.
Mitochondrial haplogroups A, B and G originated about 50,000 years ago, and bearers subsequently colonized Siberia, Korea and Japan, by about 35,000 years ago. Parts of these populations migrated to North America during the Last Glacial Maximum.
Last Glacial Maximum
Eurasia
Around 20,000 years ago, approximately 5,000 years after the Neanderthal extinction, the Last Glacial Maximum forced northern hemisphere inhabitants to migrate to several shelters (known as refugia) until the end of this period. The resulting populations are then presumed to have resided in such refuges during the LGM to ultimately reoccupy Europe where archaic historical populations are considered their descendants. The composition of European populations was later altered by further migrations, notably the Neolithic expansion from the Middle East, and still later the Chalcolithic population movements associated with Indo-European expansion. A Paleolithic site on the Yana River, Siberia, at 71°N, lies well above the Arctic Circle and dates to 27,000 radiocarbon years before present, during glacial times. This site shows that people adapted to this harsh, high-latitude, Late Pleistocene environment much earlier than previously thought.
The African Epipaleolithic Kebaran culture is believed to have reached the Near East about 18,000 years ago, introducing the bow and arrow.
Americas
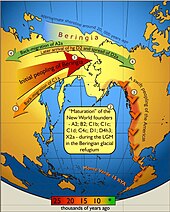
Paleo-Indians originated from Central Asia, crossing the Beringia land bridge between eastern Siberia and present-day Alaska. Humans lived throughout the Americas by the end of the last glacial period, or more specifically what is known as the late glacial maximum, no earlier than 23,000 years before present. Details of Paleo-Indian migration to and throughout the American continent, including the dates and the routes traveled, are subject to ongoing research and discussion.
The routes of migration are also debated. The traditional theory is that these early migrants moved when sea levels were significantly lowered due to the Quaternary glaciation, following herds of now-extinct pleistocene megafauna along ice-free corridors that stretched between the Laurentide and Cordilleran ice sheets. Another route proposed is that, either on foot or using primitive boats, they migrated down the Pacific coast to South America as far as Chile. Any archaeological evidence of coastal occupation during the last Ice Age would now have been covered by the sea level rise, up to a hundred metres since then. The recent finding of indigenous Australasiangenetic markers in Amazonia supports the coastal route hypothesis.
Holocene migrations

The Holocene is taken to begin 12,000 years ago, after the end of the Last Glacial Maximum. During the Holocene climatic optimum, beginning about 9,000 years ago, human populations which had been geographically confined to refugia began to migrate. By this time, most parts of the globe had been settled by H. sapiens; however, large areas that had been covered by glaciers were now re-populated.
This period sees the transition from the Mesolithic to the Neolithic stage throughout the temperate zone. The Neolithic subsequently gives way to the Bronze Age in Old Worldcultures and the gradual emergence of the historical record in the Near East and China beginning around 4,000 years ago.
Large-scale migrations of the Mesolithic to Neolithic era are thought to have given rise to the pre-modern distribution of the world's major language families such as the Niger-Congo, Nilo-Saharan, Afro-Asiatic, Uralic, Sino-Tibetan or Indo-European phyla. The speculative Nostratic theory postulates the derivation of the major language families of Eurasia (excluding Sino-Tibetan) from a single proto-languages spoken at the beginning of the Holocene period.
Eurasia
Evidence published in 2014 from genome analysis of ancient human remains suggests that the modern native populations of Europe largely descend from three distinct lineages: "Western Hunter-Gatherers", derivative of the Cro-Magnon population of Europe, Early European Farmers introduced to Europe from the Near East during the Neolithic Revolution and Ancient North Eurasians which expanded to Europe in the context of the Indo-European expansion.
The Afroasiatic Urheimat has been placed in either Africa or Asia.
Sub-Saharan Africa
The Nilotic peoples are thought to be derived from an earlier undifferentiated Eastern Sudanic unity by the 3rd millennium BC. The development of the Proto-Nilotes as a group may have been connected with their domestication of livestock. The Eastern Sudanic unity must have been considerably earlier still, perhaps around the 5th millennium BC (while the proposed Nilo-Saharan unity would date to the Upper Paleolithic about 15kya). The original locus of the early Nilotic speakers was presumably east of the Nile in what is now South Sudan. The Proto-Nilotes of the 3rd millennium BC were pastoralists, while their neighbors, the Proto-Central Sudanic peoples, were mostly agriculturalists.
The Niger-Congo phylum is thought to have emerged around 6,000 years ago in West or Central Africa. Its expansion may have been associated with the expansion of Sahel agriculture in the African Neolithic period, following the desiccation of the Sahara in c. 3900 BC. The Bantu expansion has spread the Bantu languages to Central, Eastern and Southern Africa, partly replacing the indigenous populations of these regions. Beginning about 3,000 years ago, it reached South Africa about 1,700 years ago.
Pacific

The islands of the Pacific were populated between c. 1600 BCE and 1000 CE. The Lapita people, who got their name from the archaeological site in Lapita, New Caledonia, where their characteristic pottery was first discovered, were an Austronesian-speaking people who settled in Near Oceania (notably the Bismarck Archipelago in Papua New Guinea, and the Solomon Islands) around 1500 BCE, where some intermingling with the existing Papuan population took place. Acquiring long distance voyaging skills, they ventured into 'Remote Oceania', probably settling Vanuatu and New Caledonia around 1200 BCE, then Fiji, Samoa and Tonga. By the beginning of the 1st millennium BCE, this western part of Polynesia was a loose web of thriving populations settled on the islands' coasts and living off the sea. By 0 CE Micronesia was completely colonized; tropical eastern Polynesia, including Tahiti, was probably settled by 700 CE. The last region of Polynesia to be reached was New Zealand, probably by 1300 CE.
Arctic
The last region to be permanently settled by human migrations is the Arctic.
The earliest inhabitants of North America's central and eastern Arctic are referred to as the Arctic small tool tradition (AST) and existed c. 2500 BC. AST consisted of several Paleo-Eskimo cultures, including the Independence cultures and Pre-Dorset culture.
The Inuit are the descendants of the Thule culture, which emerged from western Alaska around AD 1000 and gradually displaced the Dorset culture.
Retrieved from: https://en.wikipedia.org/wiki/Early_human_migrationsText is available under the Creative Commons Attribution-ShareAlike License